
U.S. Department of the Interior
U.S. Geological Survey
Open-File Report 2022–1084
Prepared in cooperation with the University of British Columbia, Biodiversity Research Centre
and Beaty Biodiversity Museum and Idaho State University, Department of Biological Sciences,
Fish Ecology Laboratory
Evolutionary and Ecological Connectivity in Westslope
Cutthroat Trout (Oncorhynchus clarkii lewisi) and Mountain
Whitefish (Prosopium williamsoni) in Relation to the
Potential Influences of Boundary Dam, Washington, Idaho,
and Parts of British Columbia
Evolutionary and Ecological Connectivity in
Westslope Cutthroat Trout (Oncorhynchus
clarkii lewisi) and Mountain Whitefish
(Prosopium williamsoni) in Relation to
the Potential Influences of Boundary
Dam, Washington, Idaho, and Parts of
British Columbia
By Jason B. Dunham, Eric B. Taylor, and Ernest R. Keeley
Prepared in cooperation with the University of British Columbia, Biodiversity
Research Centre and Beaty Biodiversity Museum and Idaho State University,
Department of Biological Sciences, Fish Ecology Laboratory
Open-File Report 2022–1084
U.S. Department of the Interior
U.S. Geological Survey
U.S. Geological Survey, Reston, Virginia: 2022
For more information on the USGS—the Federal source for science about the Earth, its natural and living resources,
natural hazards, and the environment—visit https://www.usgs.gov or call 1–888–ASK–USGS.
For an overview of USGS information products, including maps, imagery, and publications, visit
https://store.usgs.gov/.
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the
U.S. Government.
Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials
as noted in the text. Permission to reproduce copyrighted items must be secured from the copyright owner.
Suggested citation:
Dunham, J.B., Taylor, E.B., and Keeley, E.R., 2022, Evolutionary and ecological connectivity in westslope cutthroat
trout (Oncorhynchus clarkii lewisi) and mountain whitefish (Prosopium williamsoni) in relation to the potential
influences of Boundary Dam, Washington, Idaho, and parts of British Columbia: U.S. Geological Survey Open-File
Report 2022–1084, 22 p., https://doi.org/ 10.3133/ ofr20221084.
ISSN 2331-1258 (online)
iii
Acknowledgments
This work benefitted greatly from discussions with Ryan Simmons, Harry Rich, Al Solonsky,
and Troy Jaecks with Seattle City Light. Emily Heaston (U.S. Geological Survey, Forest and
Rangeland Ecosystem Science Center) prepared maps for this report. Reviews by Tom Quinn
(University of Washington, School of Aquatic & Fishery Sciences) and Carl Ostberg
(U.S. Geological Survey, Western Fisheries Research Center) improved an earlier draft of the
manuscript. Funding for this effort was provided by Seattle City Light.
v
Contents
Acknowledgments ........................................................................................................................................iii
Abstract ...........................................................................................................................................................1
Introduction.....................................................................................................................................................1
Part 1. Lake Pend Oreille and Pend Oreille River—Past to 2022 ...........................................................3
Overview.................................................................................................................................................3
Basin History: 15,000 Years to 2022 ....................................................................................................3
Contemporary Conditions ....................................................................................................................4
Boundary Dam and Reservoir .............................................................................................................4
Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and
Mountain Whitefish .........................................................................................................................4
Westslope Cutthroat Trout ..................................................................................................................5
Biogeography ...............................................................................................................................5
Genetic Evidence of Connectivity .............................................................................................5
Ecological Evidence of Connectivity ........................................................................................8
Environmental Conditions ...........................................................................................................9
Mountain Whitefish ............................................................................................................................12
Biogeography .............................................................................................................................12
Genetic Evidence of Connectivity ...........................................................................................12
Ecological Evidence of Connectivity ......................................................................................14
Part 3. Conclusions and Recommendations for Upstream Passage over Boundary Dam ..............15
Westslope Cutthroat Trout ................................................................................................................15
Mountain Whitefish ............................................................................................................................16
Overall Conclusions .....................................................................................................................................16
References Cited..........................................................................................................................................17
Appendix 1. Summary and Update on Connectivity for Bull Trout (Salvelinus
confluentus) in the Pend Oreille River since Dunham and Others (2014) .............................22
Figures
1. Photograph showing Boundary Dam photographed after dark, December 8, 1998 ..........2
2. Map showing the lower Clark Fork River, Lake Pend Oreille, and Pend Oreille
River with major dams indicated ................................................................................................3
3. Map showing the Pend Oreille River and associated major rivers of British
Columbia, Washington, Idaho, and Montana ...........................................................................6
4. Bar graphs showing proportional composition of samples of westslope
cutthroat trout with respect to two major genetic groups based on the single
nucleotide polymorphism analysis of Small and others .........................................................7
5. Historical, mid-century, and end-of-century predicted presence of native
westslope cutthroat trout, assuming the absence of nonnative brook trout
within the Lake Pend Oreille– Pend Oreille River system in the United States ................10
6. Pie diagrams showing genetic assignment of westslope cutthroat trout
sampled from Boundary Reservoir and Box Canyon Reservoir into major spatial
groupings .....................................................................................................................................11

vi
7. Dimensionless graph showing major genetic groups of mountain whitefish in
the Boundary Area resolved using microsatellite DNA analysis .......................................12
8. Dimensionless graph showing major genetic groups of mountain whitefish in
the Boundary Area resolved using allelic variation at microsatellite DNA loci
projected using factorial correspondence analysis .............................................................13
Conversion Factors
U.S. customary units to International System of Units
Multiply By To obtain
Length
foot (ft) 0.3048 meter (m)
mile (mi) 1.609 kilometer (km)
Flow rate
cubic foot per second (ft
3
/s) 0.02832 cubic meter per second (m
3
/s)
International System of Units to U.S. customary units
Multiply By To obtain
Length
meter (m) 3.281 foot (ft)
kilometer (km) 0.6214 mile (mi)
Area
hectare (ha) 2.471 acre
Volume
cubic meter (m
3
) 35.31 cubic foot (ft
3
)
Flow rate
cubic meter per second (m
3
/s) 35.31 cubic foot per second (ft
3
/s)
Temperature in degrees Celsius (°C) may be converted to degrees Fahrenheit (°F) as follows:
°F = (1.8 × °C) + 32.
Datums
Vertical coordinate information is referenced to the North American Vertical Datum of 1988
(NAVD 88).
Horizontal coordinate information is referenced to the North American Datum of 1983 (NAD 83).
vii
Abbreviations
WDFW Washington Department of Fish and Wildlife
SNP single nucleotide polymorphism
PIT passive integrated transponder
PoR-C Pend Oreille and Columbia Rivers

Evolutionary and Ecological Connectivity in Westslope
Cutthroat Trout (Oncorhynchus clarkii lewisi) and
Mountain Whitefish (Prosopium williamsoni) in Relation
to the Potential Influences of Boundary Dam, Washington,
Idaho, and Parts of British Columbia
By Jason B. Dunham, Eric B. Taylor, and Ernest R. Keeley
Abstract
In this report, we consider evolutionary and ecological
connectivity for westslope cutthroat trout (Oncorhynchus
clarkii lewisi) and mountain whitesh (Prosopium
williamsoni) within the Pend Oreille River in northeastern
Washington State, northern Idaho, and adjacent portions of
southeastern British Columbia, Canada. Specically, we
focused on the rationale for active translocation of individuals
of these species upstream from Boundary Dam both in
the context of natural patterns of pre-dam evolutionary
connectivity as well as preserving contemporary ecological
and evolutionary characteristics of local extant populations.
Boundary Dam impounds the Pend Oreille River (called the
Pend d’Oreille River in Canada) with the resulting reservoir
inundating two historical barriers to upstream movement
of sh (Metaline Falls and Z Canyon). Historically, it was
thought these barriers impeded the upstream movement of
westslope cutthroat trout and mountain whitesh, as well
as Pacic salmon (Oncorhynchus spp.), steelhead trout
(O. mykiss), and other resident species such as bull trout
(Salvelinus conuentus). To address connectivity, we consider
historical and contemporary processes and features. This
review includes an assessment of postglacial processes
within the Pend Oreille River and systems upstream that
include Priest Lake, Lake Pend Oreille, the Clark Fork River,
features of Boundary Reservoir and its tributaries, and areas
downstream in the Pend Oreille River such as the Salmo
River. Based on this information, we then give a more detailed
review of existing genetic and ecological data to summarize
what is known about connectivity for westslope cutthroat
trout and mountain whitesh. Our assessment of the collective
evidence leads us to conclude that moving sh upstream over
Boundary Dam is not warranted if the management objective
is to maintain natural patterns of evolutionary and ecological
connectivity or to conserve unique ecological and evolutionary
characteristics of extant local populations of these species
in the system. These ndings parallel that of a previous
analysis of bull trout. Although we were able to arrive at
well-supported conclusions in relation to Boundary Dam, we
suggest that more work on connectivity further upstream in the
Pend Oreille River would help to better understand the role of
historical processes and dams further up in the system.
Introduction
Across the United States, there are over 91,000 large
dams
1
on rivers (U.S. Army Corps of Engineers, 2018
2
).
Assessing their eects on riverine ecosystems is complicated
because dams have inuences downstream and upstream
from their location and create novel reservoir habitats
within the waters they hold back (Bellmore and others,
2019). Furthermore, the unique setting of where dams are
constructed, how they are operated, their relationships to
other barriers, and a host of other potential inuences defy
simple generalizations. Accordingly, it is important to consider
the unique context within which any given dam is located.
Although dams inuence riverine ecosystems in many ways,
one of the most fundamental involves how they inuence
evolutionary and ecological connectivity of physical and
biological systems.
Here, we consider how dams impact connectivity
for two species: westslope cutthroat trout (Oncorhynchus
clarkii lewisi) and mountain whitesh (Prosopium
williamsoni) within the Pend Oreille River in northeastern
1
The Army Corps of Engineers National Inventory of Dams includes dams
that either (1) pose a threat to human life if the dam fails, (2) pose economic
loss, environmental damage, disruption of lifeline facilities or other important
concerns, (3) equal or exceed 7.6 meters in height or 82,000 square meters of
storage, or (4) equal or exceed 1.8 meters in height and exceed 164,000 square
meters of storage.
2
Army Corps of Engineers National Inventory of Dams database can be
accessed at https:/ /nid.usace .army.mil/ #/

2 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Washington State and adjacent portions of southeastern
British Columbia, Canada (where the river is known as the
Pend d'Oreille River). The geographic focus of this report
is Boundary Dam (g. 1), which is located on the Pend
Oreille River near the U.S. - Canada border, and is one of
several dams in this watershed (g. 2). Like most salmonid
shes, westslope cutthroat trout and mountain whitesh are
inuenced by and rely on connectivity within river networks
or riverscapes (Northcote and Ennis, 1994; Neville and others,
2006a). Our specic objective in this report is to evaluate
historical and contemporary evolutionary and ecological
connectivity for westslope cutthroat trout and mountain
whitesh to address the rationale of potentially providing
upstream passage over Boundary Dam for these species.
A previous analysis (Dunham and others, 2014) examined
similar questions concerning connectivity in bull trout
(Salvelinus conuentus) in relation to Boundary Dam (app. 1).
To address our objective, Part 1 of this report begins
with a review of historical and contemporary processes
and features of the riverscape that inuence connectivity
relative to Boundary Dam within the lower Pend Oreille
system. Historically, it was thought that the upstream extent
of anadromous Pacic salmon (Oncorhynchus spp.) and
migratory rainbow trout (O. mykiss, commonly referred
to as steelhead trout) in the Pend Oreille River was just
downstream from Metaline Falls, the current approximate
location of Boundary Dam (Scholz and others, 1985). From
this foundation, in Part 2, we review existing genetic and
ecological information to evaluate what is known about
connectivity for westslope cutthroat trout and mountain
whitesh within the system. In Part 3, we assess this
evidence with respect to the evolutionary or ecological
rationale for passing these species upstream over Boundary
Dam. Throughout, we identify important uncertainties
and information gaps that could be addressed to inform
management decisions and actions to benet westslope
cutthroat trout and mountain whitesh within the area.
Figure 1. Boundary Dam (downstream view) photographed after dark, December 8, 1998 (from City of Seattle Archives, Item 126710,
Record Series 0207–01).

Part 1. Lake Pend Oreille and Pend Oreille River—Past to 2022 3
Figure 2. Lower Clark Fork River, Lake Pend Oreille, and Pend Oreille River with major dams indicated.
Part 1. Lake Pend Oreille and Pend
Oreille River—Past to 2022
Overview
The system of interest includes Lake Pend Oreille in
northern Idaho and its outlet: the Pend Oreille River (g. 2).
Lake Pend Oreille is the terminus of the Clark Fork River,
which ows westward and drains much of western Montana,
with some drainages extending north of the U.S. - Canada
border. The Pend Oreille River produces an annual average
discharge of 26,267 cubic feet per second (ft
3
/s) according
to a U.S. Geological Survey streamgage (USGS streamgage
12396500, 68 water years of record) located near Box
Canyon Dam, which is 55.2 kilometers (km) upstream from
the conuence of the river with the mainstem of the upper
Columbia River in southeastern British Columbia. From Lake
Pend Oreille to the Columbia River, the Pend Oreille River
ows for 210 km. A major Pend Oreille River tributary near
the outlet of Lake Pend Oreille and just upstream from Albeni
Falls (
g. 2) is the Priest River, which drains Priest Lake and
discharges an annual average of 1,705 ft
3
/s (USGS streamgage
12395000, 71 water years of record) into the system.
Basin History: 15,000 Years to 2022
Between approximately 13,000–15,000 years ago, most
of the Pend Oreille River was beneath the Purcell Lobe of the
Cordilleran Ice Sheet. The Purcell Lobe covered the bottom
of present-day Lake Pend Oreille and dammed the Clark Fork
River, forming glacial Lake Missoula. This massive water
body (over 320 km long and 610 meters [m] deep) was the
largest ice-dammed lake known to have existed (O'Connor
and others, 2020). The waters behind the ice dam were
released with catastrophic ooding downstream extending
across eastern Washington and occurring several times as the
dam re-formed and failed in relation to uctuations in the Ice
Sheet. Following glacial retreat about 11,000 years ago, Lake
Pend Oreille and the Pend Oreille River began to assume their
present forms (McPhail and Lindsey, 1986; Pielou, 1991; Link
and Keeley, 2018; O'Connor and others, 2020).
4 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
With the river and lake system formed, native shes
gained access to the area from glacial refugia located upstream
and downstream from the inuence of the Cordilleran
Ice Sheet (McPhail and Lindsey, 1986). Although native
salmonids such as bull trout, mountain whitesh, and
westslope cutthroat trout were able to colonize throughout
the system, anadromous Pacic salmon and rainbow trout in
the Pend Oreille River are thought to have been limited to
areas below the hydraulic barriers posed by Z Canyon and
Metaline Falls (near the present site of Boundary Dam, see
below; Scholz and others, 1985). Historical accounts indicate
that anadromous shes were limited by the falls and cascades
in the lower Pend Oreille River. There is some uncertainty,
however, if any single waterfall completely blocked the
passage of salmon and steelhead trout in all years, at least
up to Albeni Falls (Scholz and others, 1985). Within most
tributaries to Boundary Reservoir (the impoundment above
Boundary Dam), natural waterfalls and steep cascades also
likely acted to restrict the movement of native shes and this
continued with the construction of Boundary Dam. Beyond
migratory shes, accounts of resident sh consumption by the
Kalispel tribe indicate signicant whitesh, char (Salvelinus,
sp.), trout, and sucker (Catostomidae) populations in the Pend
Oreille River (Scholz and others, 1985).
Contemporary Conditions
Today the Pend Oreille River is inuenced by a series
of large dams, beginning with Cabinet Gorge Dam, which
is located just upstream from Lake Pend Oreille on the
Clark Fork River and controls ows into the lake (g. 2).
Downstream from Lake Pend Oreille, major dams include
the Priest Lake Outlet Dam on the Priest River, and on the
Pend Oreille River, three dams in the United States (Albeni
Falls, Box Canyon, and Boundary dams) as well as two in
British Columbia (Seven Mile and Waneta dams). There are
a few dams on key tributaries, including Mill Pond Dam
(removed in 2017) and Sullivan Lake Dam in the Sullivan
Creek basin. Numerous stream-road crossings (for example,
culverts) and other human-built structures are present and
sometimes hinder or prevent sh passage. A host of nonnative
aquatic animals, ranging from mysid shrimp (Mysis spp.)
to northern pike (Esox lucius) and dozens of other species
have become established in the Pend Oreille system from
intentional human introductions and subsequent dispersal.
Active suppression eorts are occurring to control nonnative
lake trout (S. namaycush; Hansen and others, 2010), brook
trout (S. fontinalis; Dunham and others, 2002), and northern
pike (Carim and others, 2019) to minimize their inuences
on native species and ecosystem processes. In short, ow
regulation, introductions and invasions of nonnative species,
and other changes wrought by humans to Lake Pend Oreille
and the Pend Oreille River have fundamentally transformed
the ecosystem. Despite such transformations, native species
persist throughout much of the system.
Boundary Dam and Reservoir
This report focuses on the specic question of how
Boundary Dam inuences connectivity for westslope cutthroat
trout and mountain whitesh. Accordingly, it is important
to describe a few essential details of the dam, the associated
reservoir, and tributaries. The salient characteristics of
Boundary Dam were summarized from the City of Seattle's
description of the project (Frantilla, 2022, http s://www.se
attle.gov/ cityarchives/ exhibits- and- education/ online- exhibits/
boundary- dam). Construction of Boundary Dam was
completed in 1967, stands at just over 100 m tall, and can
produce 127 million kilowatts of electricity on an annual
basis. The impoundment upstream from Boundary Dam, the
Boundary Reservoir, has a maximum depth of about 82 m, a
maximum surface area of about 726 hectares (ha), and extends
for 28 km upstream to the base of Box Canyon Dam. Water
ow through the reservoir is relatively fast, with a residence
time that ranges between 2 and 4 days (Pickett, 2004).
Correspondingly, temperatures are fairly consistent up and
downstream, and the reservoir is not thermally stratied at any
time of the year (Seattle City Light [SCL], 2006).
Seattle City Light’s (2006) assessment listed 28
tributaries to Boundary Reservoir that oer a total of nearly 25
km of stream distance that is accessible to upstream-migrating
salmonids (Washington Department of Fish and Wildlife
[WDFW], 2020). For individual tributaries, accessible stream
distance ranges from 0 km (barrier falls at the conuence) to
potentially 32 km in Sullivan Creek after the 2017 removal
of Mill Pond Dam (Federal Energy Regulatory Commission
[FERC], 2011). Despite the removal of this dam, however, a
seasonal velocity barrier approximately 1 km from the mouth
of Sullivan Creek remains (Powers, 2008).
Part 2. Evolutionary and Ecological
Connectivity for Westslope Cutthroat
Trout and Mountain Whitefish
In this section, we review evidence describing historical
and contemporary patterns of connectivity for westslope
cutthroat trout and mountain whitesh. Although the focus
of our review is on upstream passage over Boundary Dam,
we review salient local and regional factors inuencing
connectivity. In Part 3 below, we rely on the body of evidence
reviewed here to draw conclusions regarding the value of
providing passage for these species upstream over Boundary
Dam. In addition to considering these species, we briey
revisit the case for bull trout (Dunham and others, 2014; app.
1) to provide a more complete and updated review of upstream
passage over Boundary Dam.
Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and Mountain Whitefish 5
Westslope Cutthroat Trout
Biogeography
The Pend Oreille River represents a transitional region
in the distribution between two native trout species in western
North America: rainbow trout and westslope cutthroat trout.
The natural biogeographic history of the boundary area is
thought to have been strongly inuenced by the various
Pleistocene glaciations (roughly 2.8 million to 18,000 before
present [B.P.]) that covered vast swaths of land and had
dramatic inuences on streamow and watershed connectivity
over time (McPhail and Lindsey, 1986; Pielou, 1991).
Although the exact biogeographic history of the area cannot be
fully reconstructed today, recolonization of ice-covered areas
by sh species must have occurred following glacial retreat.
Areas outside of the extent of glaciation may have also been
greatly aected by historical oods, elevated stream ows,
and interconnection of watersheds, allowing dispersal of sh
species over areas now presently isolated from each other.
Based on historical records and observations from the late
1800s, Z Canyon (immediately upstream from the present-day
location of Boundary Dam and Metaline Falls on the Pend
Oreille River 15 km farther upstream) appears to have limited
or at least impaired upstream movement of anadromous shes,
which would likely have included steelhead trout (Scholz and
others, 1985; Nellestijn and Ells, 2008).
Contemporary evidence for connectivity of westslope
cutthroat trout relative to Boundary Dam is available from
genetic and distribution data as well as from studies of sh
movements and demographic characteristics of populations
within tributary systems. Arguably, the most comprehensive
description of trout distribution patterns in North America
was written by R.J. Behnke (1992, 2002). He described the
native distribution of westslope cutthroat trout to follow the
west side of Kootenay Lake in British Columbia and south
into Idaho but outside most of the Priest Lake watershed
and the downstream sections of the Pend Oreille River
(g. 3). Although McPhail (2007) recorded observations of
westslope cutthroat trout in the watersheds noted by Behnke
(1992, 2002) around the Kootenay River of southeast British
Columbia, McPhail (2007) also reported that westslope
cutthroat trout populations occur in the Salmo River
downstream from Boundary Dam and in other tributaries
of the upper Columbia River outside of the continuous
distribution estimated by Behnke (1992, 2002). More recent
sampling in the boundary area established that westslope
cutthroat trout populations exist in tributaries of the Pend
Oreille River and in the Priest Lake watershed (g. 3; Small
and others, 2020).
Genetic Evidence of Connectivity
As with bull trout (Dunham and others, 2014), the
relationship of westslope cutthroat trout in the Salmo
River to populations upstream from Boundary Dam and
historical barriers to upstream passage (in other words,
Metaline Falls and Z Canyon) in the Pend Oreille system can
provide important clues about historical and contemporary
connectivity. Overall, there is little evidence that suggests
any appreciable connectivity between native westslope
cutthroat trout below Boundary Dam and upstream areas. For
instance, genetic analysis of westslope cutthroat trout began
with an analysis of 10 microsatellite DNA loci in 2007 by the
Washington Department of Fish and Wildlife (Washington
Department of Fish and Wildlife [WDFW], 2020; Small and
others, 2007). This study included 77 localities within the
Pend Oreille and Priest Lake watersheds as well as several
groups of hatchery sh used in various stocking programs
(westslope cutthroat trout, Yellowstone cutthroat trout,
and rainbow trout). The Salmo River collection included
samples from near the conuence with the Pend Oreille River
(Swift Creek, about 12 km upstream) and a small lake in the
headwaters (Waldie Lake). Unfortunately, owing to variable
and uncertain scoring of microsatellite loci for the Salmo
River samples, little insight was gained on the status of these
sh (M. Small, oral comm., May 2020). The few analyses
that were completed indicated that the Waldie Lake and Swift
Creek samples were quite distinct from one another (both were
more similar to samples outside the Salmo River drainage than
they were to each other) as well as from other sh from the
Pend Oreille study area.
Furthermore, Small and others (2017) reported the results
of analyses of about 1,850 samples, including sh from ve
sites within the Salmo River watershed, using a panel of 215
single nucleotide polymorphism (SNP) loci. In this analysis,
sh were rst screened for hybridization with rainbow trout
using four loci that were diagnostic for dierences between
the species. Results indicated that 29 percent of sh sampled
from the Salmo River sites (range 3–68 percent) were
interspecic hybrids between rainbow trout and cutthroat trout
(table 3 in Small and others, 2017). Even the most upstream
sample sites (Crutch and Watch Creeks) showed some
evidence of introgression with rainbow trout although at much
lower levels (3–6 percent) than in downstream reaches of the
Salmo River system. Of the remaining Salmo River sh that
were deemed to be westslope cutthroat trout from four sites
(two were pooled as they were so similar to one another), all
showed very high genetic anity to the King's Lake (Priest
Lake drainage origin) hatchery samples of westslope cutthroat
trout (g. 4). These sh showed a similar “hatchery genetic
signal” to sh from other localities within the Pend Oreille
drainage that have been subject to hatchery supplementation
(for example, Sweet, Slate, Lunch, Lost, and Harvey Creeks).
The U.S. part of the Salmo River has also received at least
some supplementation of westslope cutthroat trout using the
King's Lake hatchery source (Small and others, 2017).

6 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Figure 3. Pend Oreille River and associated major rivers (blue lines) of British Columbia, Washington, Idaho, and Montana. Brown
polygon represents the estimated western extent of the native distribution of westslope cutthroat trout (Oncorhynchus clarkii lewisi) in
southeastern British Columbia (McPhail, 2007). The light brown polygon is the estimated extent of westslope cutthroat trout by Behnke
(1992, 2002). Hatched area indicates the estimated overlap in range between McPhail (2007) and Behnke (1992, 2002). Red circles
represent sampling locations for westslope cutthroat trout populations reported in Small and others (2020). Black crosses indicate the
location of major dams in the region. Blue crosses indicate the location of known waterfalls (World Waterfall Database, 2022, https://w
ww.worldwa terfalldat abase.com/ ).

Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and Mountain Whitefish 7
KLH
SR
16Slumber
99KingLk
12KingBY11
12KingBY12
02Sweet
12Sweet1
12Sweet2
14Watch
04Lost
14Crutch
14SSalR5UpST
12Uncas
99Slate
12Slate1
12Slate2
13Slate2.6
12Slate3
12SFSlate
12Styx
02Lunch
12Lunch1
12Lunch2
12Pewee
12MFFlume
11Deem0.6
12Deem3.6
12DeemUp
11_13Gypsy1
11GypsyNE
11_13GypsyWB
11_12John
11Kinyon0.8
11Kinyon0.6
11Leola0.7
11_13LeolaNE
11Leola1.6
13Leola0
13Leola1.6
13LeolaNW
99NFSull
03NFSull
12NFSull1
13NFSull1
12NFSull2
11Pass0.5
12PassUp
11_12RainyUp
11Rainy0.2
12RainLo
13Rain
Lo
13Smart
11Stony2.2
03SullLk
12Harvey1
02Harvey
12Noisy
99UpSull
14SullCr
12Sull11
12Sull14.5
13Sull16.5
11Sull16.4
13Sull18.5
11Sull19
11Sull20.7
13Sull21
KingsLk
Sullivan
Figure 4. Bar graphs showing proportional composition of samples of westslope cutthroat trout (Oncorhynchus clarkii lewisi) with
respect to two major genetic groups (blue and red shading) based on the single nucleotide polymorphism analysis of Small and others
(2017). Each bar represents the mean proportional contribution (0.0 to 1.0) of fish from each locality across multiple admixture analyses.
Fish from the King's Lake hatchery (KLH, three samples) are predominantly composed of the blue genetic group. Fish from localities
within the Salmo River (SR, three samples) are also predominantly composed of the blue group, as are some tributaries of the Sullivan
Creek (“Sull”) drainage and the Pend Oreille River upstream from Boundary Dam that have been subject to hatchery supplementation
with King's Lake hatchery westslope cutthroat trout (indicated by blue stars; for example, Harvey, Lunch, Lost, Slate, North Fork Sullivan
(NFSull) Creeks, see Small and others 2017, table 1 and fig. 2). The red genetic group is thought to be characteristic of native fish from
the Sullivan Creek watershed.
8 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Next, Small and others (2019a) reported the results
of genetic tests that used a baseline of SNP information on
potential source populations to genetically assign sh (N = 16)
sampled from various non-tributary localities to these potential
source populations. One sh was sampled from the Boundary
Dam tailrace, and it was estimated to have originated from
either Watch Creek (a tributary of the uppermost reaches of
the Salmo River in Washington) or Slate Creek, a tributary of
the Pend Oreille about 10 km upstream from the Boundary
Dam. Consequently, identication of this tailrace westslope
cutthoat trout was ambiguous relative to whether it may have
originated from upstream or downstream from the dam site.
The remaining 15 sh were all sampled from areas upstream
from the dam and were identied as either rainbow trout
(N = 1), westslope cutthroat trout x rainbow trout hybrids
(N = 3 sh of varying levels of admixture between the two
species), King’s Lake hatchery westslope cutthroat trout (N =
1), either native or hatchery sh (N = 2), or native sh from
localities upstream from the dam, including the Sullivan Creek
drainage (N = 8).
Finally, A similar analysis was conducted by Small and
others (2020) for a sample of 47 sh, all but seven of which
were sampled in river sections between Boundary and Box
Canyon dams. The largest percentage of sh were either
introgressed (up to 75 percent introgression) with rainbow
trout (19 of 47 sh, 40.4 percent), or were assigned to the
King’s Lake westslope cutthroat trout hatchery stock (13 of 47
sh, 27.7 percent). Two more sh were identied as rainbow
trout. Of the remaining 13 sh, a single individual was
considered to have originated from the Salmo River or King's
Lake hatchery, and another was assigned to the Salmo River or
Granite Creek (one of the original brood sources for the King's
Lake hatchery strain; Small and others, 2007, 2020), three sh
were assigned as Salmo River watershed westslope cutthroat
trout, and eight sh were assigned to various localities
between Boundary and Box Canyon dams. In summary, of
40 sh sampled above the Boundary Dam, only one was
unequivocally assigned to the Salmo River drainage (and
thus suggests some historical connectivity between the Salmo
River and the Pend Oreille River upstream of Metaline Falls).
Additionally, of the seven sh sampled below Boundary Dam,
only one could be unequivocally assigned to a site upstream
from the dam (to Lunch Creek), suggesting either entrainment
through the dam or some historical connectivity through
Metaline Falls.
In sum, the extensive genetic sampling and analysis
by WDFW strongly suggest that westslope cutthroat trout
sampled from the Salmo River (and the few sh sampled in
the tailrace sections and genetically assigned to the Salmo
River baseline) are either: (1) hybrids between rainbow
trout and westslope cutthroat trout or, (2) are genetically
indistinguishable from the King's Lake hatchery strain. Small
and others (2017) concluded that Salmo River westslope
cutthroat trout included in their study were either a population
founded from hatchery plantings or a native population
that has been substantially inuenced by introgression with
hatchery sh. The likelihood of a hatchery origin of westslope
cutthroat trout in at least the mainstem reaches of lower
Salmo River is considered to be high by sheries workers in
the Nelson, BC, area (J. Baxter, FortisBC, written commun.
with Eric Taylor, May 2020). Furthermore, consultation with
local authorities could not conrm any native populations
of westslope cutthroat trout between the conuence of the
Salmo and Pend Oreille Rivers and the conuence of the latter
with the Columbia River (C. Lee, WDFW, Spokane, and T.
Anderson, Kalispel Tribe Fish and Wildlife Dept., written
commun. with Eric Taylor, May 2020).
The lack of signicant historical connectivity between
populations of westslope cutthroat trout downstream and
upstream from Boundary Dam is consistent with conclusions
for bull trout (Dunham and others, 2014). Finally, Chinook
salmon (O. tshawytscha) and steelhead trout have migrated
to the conuence of the Salmo River (formerly referred to
as the “Salmon” River) and the Pend Oreille River as well
as upstream in the Salmo River, but Metaline Falls and Z
Canyon probably represented barriers to further upstream
migration for anadromous sh in the Pend Oreille River, and
thus almost surely served as barrier to upstream migration for
westslope cutthroat trout and mountain whitesh (Nellestijn
and Ells, 2008).
Ecological Evidence of Connectivity
Inferences from molecular markers can provide important
insights into potential patterns of historical and contemporary
connectivity (Neville and others, 2006a, b). Many forms
of connectivity cannot be easily evaluated using molecular
markers alone, however. Complex patterns of sh movement
and consequences for growth, survival, and reproduction
require tracking of tagged sh or indirect methods (for
example, observational or modeling studies) to evaluate the
importance of connectivity in riverscapes (Young, 1996; Falke
and Fausch, 2010).
Direct evidence of movement of westslope cutthroat
trout in Boundary Reservoir and its tributaries comes from
tagging and tracking studies. In the reservoir and tailrace,
56 westslope cutthroat trout were tagged and tracked using
acoustic telemetry between 2016 and 2019 (Hydroacoustics
Technology, Inc., 2018, 2020). Tagged sh were tracked
between the tailrace of Boundary Dam and downstream
areas, as well as in Boundary Reservoir up to the base of
Box Canyon Dam. Available reports mention that the length
and mass of tagged individuals were recorded, but were not
reported, so eects of size on movement cannot be evaluated
(for example Monnot and others, 2008). These sh were
captured over the course of 24 electroshing events, each of
which involved at least 1 day of eort and yielded a catch
rate of less than two sh per event. Interestingly, over the
course of the 3-year study and sampling eorts,10 of the 56
(18 percent) tagged sh were recaptured. Of all sh tagged
and tracked over 3 years, eight individuals (14 percent) were
entrained downstream through Boundary Dam. The average
Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and Mountain Whitefish 9
size of westslope cutthroat trout entrained (x
̅
= 286 millimeter
fork length; standard deviation [SD]= 61) were statistically
indistinguishable to those that were not entrained (x
̅
= 310
mm fork length; SD = 66; Hydroacoustics Technology
Incorporated, 2018, 2020, 2021).
Overall, studies of westslope cutthroat trout indicate
that very few individuals currently occupy Boundary
Reservoir, though precise density estimates are not available.
Furthermore, only a few of these individuals were entrained
downstream through Boundary Dam (14 percent). Low
numbers of tagged and tracked individuals, uncertainties
about size-related movement patterns, and lack of tracking of
individuals from the reservoir into tributaries limit what can be
concluded from this work in terms of patterns of connectivity
beyond the simple question of entrainment.
Tagging of westslope cutthroat trout in tributaries to
Boundary Reservoir has involved considerably more locations
and individuals relative to work in the main-stem river
(West Fork Environmental, 2020). Tributary studies have
used Passive Integrated Transponders (PIT tags), and results
from work conducted from 2012 to 2019 were available for
this report. Of the 28 tributaries identied by Seattle City
Light (2006), tagging of individual westslope cutthroat
trout has been conducted in Peewee, Slate, Sullivan, Flume,
Pocahontas, and Sweet-Lunch Creeks. It is important to note
these tributaries have complete or partial natural barriers to
upstream movement of sh that occur near their conuences
with Boundary Reservoir, and that PIT tagging has occurred
upstream from these barriers. Over 8 years (2012–19), a
total of 13,975 westslope cutthroat trout were PIT-tagged.
Individuals were recaptured by electroshing at locations
distributed across tributaries. Between 2014 and 2019,
numerous 200 m locations have been annually sampled 1–6
times by single-pass electroshing. The total number of sites
sampled across all years was not reported, but in 2018, for
example, eorts included 46 sites sampled in previous years
and 32 new sites located next to previously sampled sites.
Based on existing tagging and electroshing eorts
and the distribution of sites sampled in space and time, few
westslope cutthroat trout were recaptured within or beyond
the site of initial capture. Recaptured individuals in 2019
represented sh that were tagged in previous years, including
71, 19, 6, 3, and 1 percent for sh initially PIT-tagged in
2018, 2017, 2016, 2015, and 2014, respectively. A total of
six PIT-tag arrays are being used for the detection of tagged
individuals as they swim past the antennas, including ve
in Sullivan Creek and one in Slate Creek. All but one of the
PIT array locations include three channel-spanning arrays to
ascertain the directional movement of detected individuals.
Since the antennas were installed in 2014 and 2015, the PIT
tag arrays have recorded a total of 516,602 detections from
486 unique sh. This low number of unique sh suggests that
movement across PIT arrays is generally infrequent and that
tagged sh reside primarily within tributaries (Lukacs and
Landguth, 2020). While data from the PIT arrays suggests
very few individuals leave Sullivan Creek for Boundary
Reservoir, there have been considerable challenges with
maintaining the most downstream antenna near the outlet of
Sullivan Creek, adding an unknown degree of uncertainty to
these ndings (Lukacs and Landguth, 2020). Overall, captured
sh (tagged and untagged) were relatively small (43–228 mm,
fork length) but is consistent with sizes that may be expected
for small streams. Growth rates based on recaptured sh did
not exhibit any obvious outlying values or bi-modality as may
be expected of a population composed of a mixture of resident
and migratory individuals (Kendall and others, 2015).
Environmental Conditions
Current conditions within Boundary Reservoir are
marginal for supporting westslope cutthroat trout due in part
to warm seasonal water temperatures. Consistently colder
water temperatures are available in tributary systems feeding
Boundary Reservoir and at tributary mouths. A recent report
(Environmental Science Associates [ESA], 2019) documenting
seasonal temperatures within Boundary Reservoir indicated
extended seasonal (July–Sept) occurrences of temperatures
above 20 °Celsius (C) at all locations sampled. Temperatures
in the Boundary Reservoir forebay recorded at 10 m depth
exceeded 20 ° C from July 10th through September 14th,
2019, with a maximum of 23 °C. Temperatures recorded
at 60 m were similar, reecting isothermal conditions that
should be expected in a system with water discharge that has
a short residence time. Small, coldwater refuges at Boundary
tributary mouths (H. Rich and R. Simmons, Seattle City
Light, oral communication, April 2021) are not apparent in
that dataset. Similarly, some locations of the Pend Oreille
River upstream from Box Canyon Dam, have colder patches
or thermal refuges near tributary conuences. It is not clear,
however, if the use of these cold refuges can be expected to
improve conditions for coldwater sh (for example, mountain
whitesh, westslope cutthroat trout, bull trout) as the benets
of thermal refuge use depend on many contingencies such
as other measures of habitat quality, including vulnerability
to predators, lack of feeding opportunities (Snyder and
others, 2020), poor water quality (for example, low dissolved
oxygen), or possibly the presence of other nonnative species,
such as brook trout.
Although temperatures within the Boundary Reservoir
exceed levels considered suitable for coldwater taxa such as
salmonids (Richter and Kolmes, 2005), tributaries within the
system are a potential source of cold-water habitat. Modeled
presence of cold-water species such as westslope cutthroat
trout is predicted to be high in many locations, particularly
within Sullivan Creek (Isaak and others, 2016; g. 5).
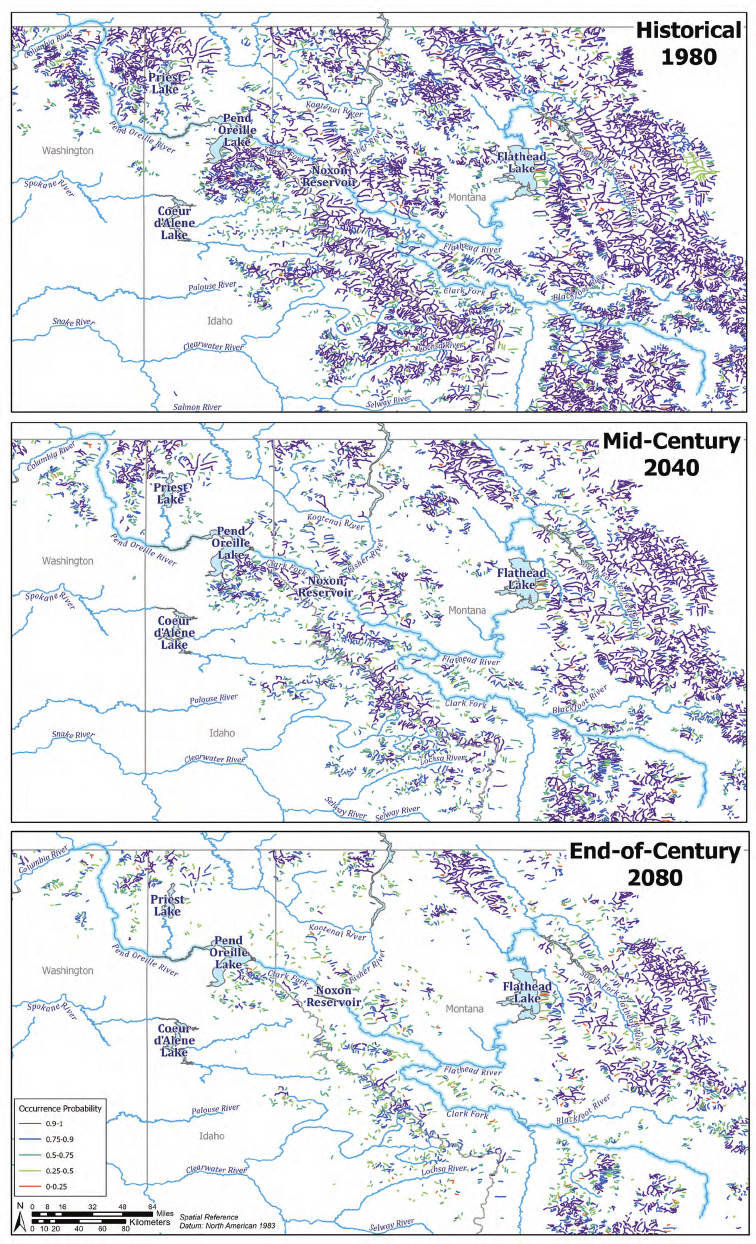
10 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Figure 5. Historical, mid-century, and end-of-century (based on climate change scenarios in
1980, 2040, and 2080) predicted presence of native westslope cutthroat trout (Oncorhynchus clarkii
lewisi), assuming the absence of nonnative brook trout (S. fontinalis) within the Lake Pend Oreille–
Pend Oreille River system (fig. 2) in the United States (from Isaak and others, 2016).

Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and Mountain Whitefish 11
Colder tributaries to Boundary Reservoir could act as
thermal refuges if they are accessible. In the largest system,
Sullivan Creek, a series of chutes and cascades begin about 1
km upstream from the conuence with Boundary Reservoir.
Such conditions are considered to pose a barrier to upstream
passage of bull trout less than 457 mm when ows exceed
about 3 m
3
/s Existing stream discharge records (USGS gage
12398000) indicate that ows often exceed those levels during
much of the year, but larger sh may be able to move upstream
into Sullivan Creek as ows decline in late summer through
fall. It is worth noting that none of the 16,980 westslope
cutthroat trout captured in tributary surveys from 2012 to
2018 were larger than 350 mm (West Fork Environmental,
2020). Sizes of individuals tracked using acoustic telemetry
in Boundary Reservoir averaged 276 mm (SD 47.3) with a
maximum length of 395 mm. Access to other tributaries of
Boundary Reservoir with available cold water is not possible
due to the presence of natural barriers near the conuences of
Peewee, Slate, Sweet, Flume, and Sand Creeks.
Before large dams were constructed in the Pend Oreille
system, it is possible that westslope cutthroat trout exhibited
migratory patterns that were similar to those exhibited by
bull trout in the system (DuPont and others, 2007, Dunham
and others, 2014). Historical patterns of movement by bull
trout included a strong inuence of seasonal emigration of
individuals from systems upstream from the Pend Oreille
River, with some of these individuals passing downstream
over the barriers of Z Canyon and Metaline Falls. There is
little doubt that the numbers of westslope cutthroat trout from
the Priest Lake, Lake Pend Oreille, and Clark Fork River
systems could also have produced individuals that migrated
downstream seasonally into the Pend Oreille River. Such
individuals would, however, have been unable to historically
access downstream tributaries between present-day Boundary
and Box Canyon dams due to natural barriers near their
conuences with the Pend Oreille River. Furthermore, the
presence of such migratory individuals would be limited to the
section of the river (which is now part of Boundary Reservoir)
that is located upstream from Metaline Falls (just downstream
from the conuence of Sullivan Creek). Individuals moving
downstream from Metaline Falls would not have been able to
return upstream.
Today, westslope cutthroat trout in Boundary Reservoir
represent a mixture of origins, including individuals from
populations upstream from Box Canyon Dam (see “Genetic
Evidence of Connectivity” above, and g. 6). Of 40 westslope
cutthroat trout sampled for n tissues in Boundary Reservoir
during 2018–19, the proportion of individuals genetically
assigned to populations upstream in Box Canyon Reservoir or
Priest River tributaries (35 percent; n = 14) was double that
assigned to Boundary Reservoir tributaries (15 percent, n = 6;
g. 6; Small and others, 2020). The proportion of individuals
assigned to the King's Lake Hatchery was even higher (40
percent; n = 16) in Boundary Reservoir. A similar genetic
analysis of westslope cutthroat trout in Box Canyon Reservoir
(n = 202) showed only 2 percent (n = 3) assigning to King's
Lake Hatchery (g. 6; Small and others, 2020).
Figure 6. Genetic assignment of westslope cutthroat trout (Oncorhynchus clarkii lewisi) sampled from Boundary Reservoir (n = 40;
left panel) and Box Canyon Reservoir (n = 202; right panel) into major spatial groupings (Boundary Reservoir tributaries, Box Canyon
Reservoir tributaries, Priest River tributaries, King's Lake Hatchery, and phenotypic westslope cutthroat trout that were assigned as
rainbow trout (Oncorhynchus mykiss); Small and others 2020).
12 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Mountain Whitefish
Biogeography
Mountain whitesh is one of the most widely distributed
yet least studied inland salmonid shes in western North
America. The species occurs south from the Lahontan and
Bonneville basins within the hydrographic Great Basin
desert into the Columbia River basin, upper Missouri River
and in other inland and coastal tributary streams north to
the Mackenzie River in the Northwest Territories, Canada
(Warren and Burr, 2014). Its widespread distribution suggests
a complex historical biogeography, but this has not been
well studied (Miller, 2006). Unlike westslope cutthroat trout
and other salmonids, mountain whitesh have not been
widely introduced by humans. Thus, indirect inferences
from molecular markers are not as subject to biases from this
potential inuence.
Genetic Evidence of Connectivity
Limited work has been conducted on mountain whitesh
in the Pend Oreille River. The most salient data are those
of Small and others (2019b), who examined variation at 15
microsatellite DNA loci in 163 individuals. The samples
ranged from one sh collected in the Boundary Dam tailrace
to sh from the former Mill Pond Dam area of the Sullivan
Creek watershed, sh from Sullivan Creek itself, and sh
from a range of localities between Boundary Dam forebay and
Albeni Falls.
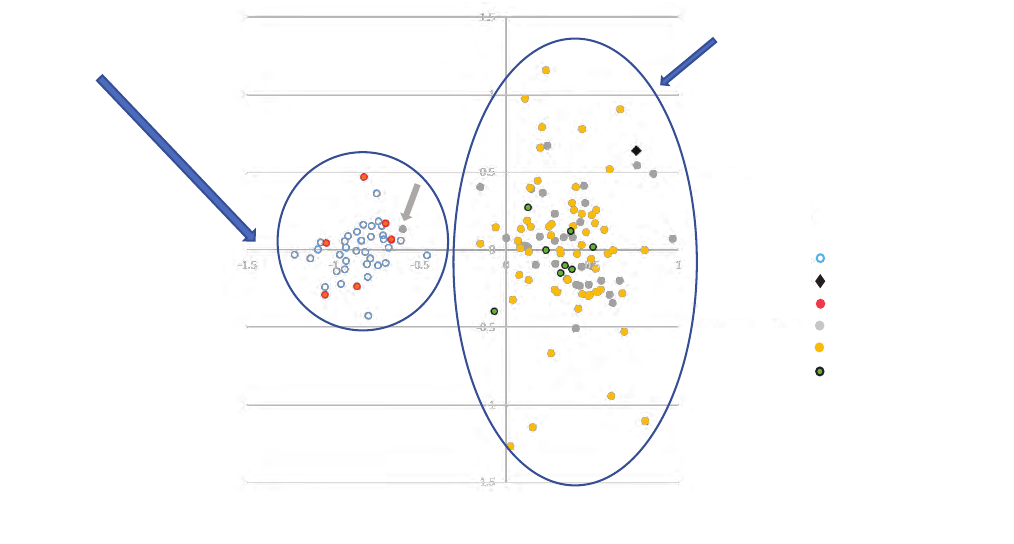
This analysis showed a major dierence between samples
in the Mill Pond/Sullivan Creek area and those collected
between Boundary Dam and Albeni Falls (g. 7). The one
sample collected in the Boundary Dam tailrace was very
similar to the sh collected from the Boundary Reservoir to
Albeni Falls area, suggesting that it originated in this area and
was entrained downstream through the dam. Furthermore,
one sh collected in the Pend Oreille River upstream from
Boundary Dam appeared to originate above the former site of
Mill Pond Dam (g. 7, grey symbol indicted by grey arrow).
By contrast, there were no dierences found among samples
collected between Boundary Dam to Albeni Falls, nor between
sh from the Mill Pond area and upper Sullivan Creek
(although sample sizes were very small in the latter group).
This analysis was augmented by further work on 41
Mountain whitesh collected from Sullivan Lake and ve
sh collected from the Pend Oreille River between Boundary
Dam and Box Canyon Dam (Small and others, 2020).
The Sullivan Lake sh were strikingly distinct from those
Mill Pond/upper Sullivan Creek
Pend Oreille River
mainstem (Albeni Falls
downstream to
Metaline Falls),
reservoir tributaries,
tailrace of Boundary
Dam
14Mill
14Boundary Tailrace
17Sullivan
12PendOreille_12AK
12PendOreille_12DF
12SweetCr_12HH
Individual site code
s
Figure 7. Major genetic groups of mountain whitefish (Prosopium williamsoni) in the Boundary Area resolved using microsatellite DNA
analysis. Samples are projected in factorial correspondence analysis space and the ellipses denote two genetic groups of mountain
whitefish: Mill Pond/upper Sullivan Creek and Pend Oreille main stem and tributaries, and membership of individual fish within those
two groups. Modified from Small and others (2019b). The grey closed symbol within the left-most ellipse represents a fish sampled from
the Boundary Reservoir and the black solid diamond within the right-most ellipse represents a fish sampled from the Boundary Dam
tailrace.
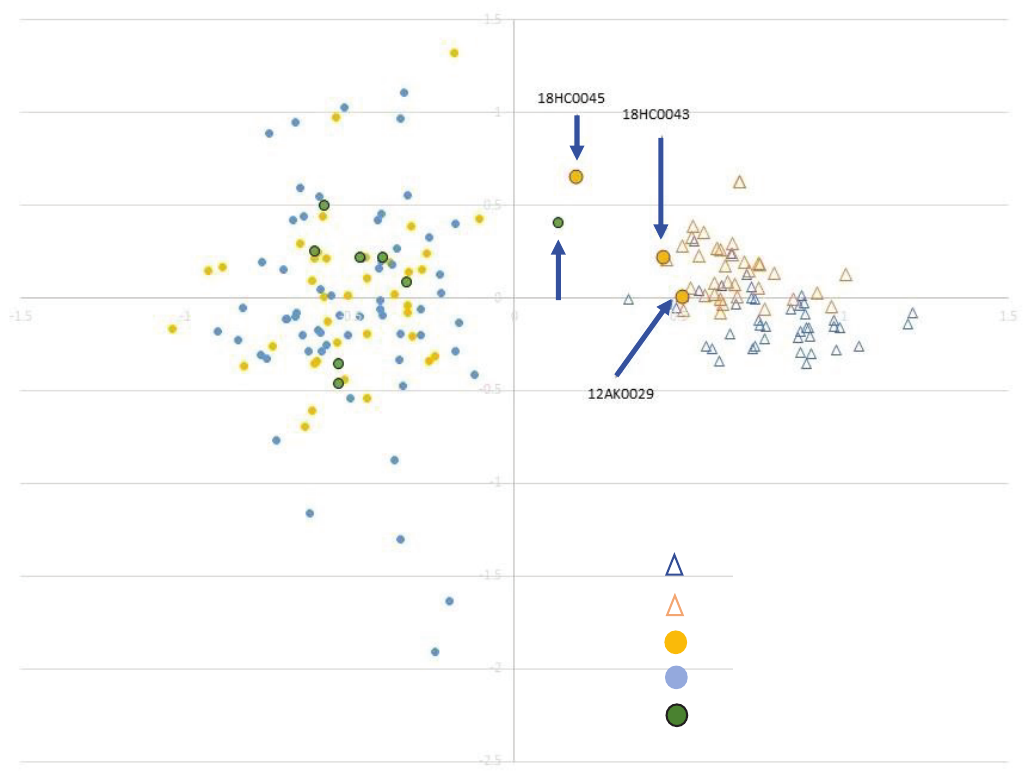
Part 2. Evolutionary and Ecological Connectivity for Westslope Cutthroat Trout and Mountain Whitefish 13
collected from the Pend Oreille River (F
ST
ranged from 0.10
to 0.11) but were only modestly (but signicantly) distinct
from Mill Pond Mountain whitesh (F
ST
= 0.025, g. 8).
Despite these average dierences among the dierent groups
of mountain whitesh, three sh collected in the mainstem
Pend Oreille River (between upstream from Boundary Dam
and downstream from Box Canyon Dam) were genetically
similar to sh from the Sullivan Lake/Mill Pond group and
remarkably similar to the Mill Pond sh (g. 8). These data
suggest the occasional downstream movement of Sullivan
Lake/Mill Pond sh into the area between Boundary Dam and
Box Canyon Dam. Notwithstanding occasional downstream
movements, the dierence between the Mill Pond/Sullivan
Creek/Sullivan Lake samples and the Boundary Dam/Albeni
Falls samples suggests that there has been restricted movement
of sh between these areas, even though the Sullivan Creek
watershed enters the Pend Oreille River above Metaline Falls.
Alternatively, it is possible that the distinction between areas
could be an artifact of the isolation created by the Mill Pond
Dam that was removed in 2017. This, however, seems unlikely
given that there was a natural water velocity barrier in Sullivan
Creek where the Mill Pond Dam was located and another
at the outlet of Sullivan Lake (Small and others, 2020).
Furthermore, despite smaller sample sizes, the Mill Pond/
Sullivan Creek/Sullivan Lake samples contained ten alleles
(at low to moderate frequencies of 1–12 percent) not observed
in the Boundary Dam/Albeni Falls samples. This observation
suggests the Mill Pond/Sullivan Creek/Sullivan Lake sh
have been isolated long enough from the Pend Oreille sh
for these novel mutations to accumulate. Furthermore, strong
genetic drift induced by isolation from Mill Pond Dam
construction would be expected to eliminate rare, unique
16_17_18SullLk
14Mill
12PendOrR
12BoxCany
12HH_Sweet
Individual site codes
1.5
1.0
0.5
0
0 1.51.00.5−1.5 −1.0 −0.5
0.5
1.0
1.5
2.0
2.5
Figure 8. Major genetic groups of mountain whitefish (Prosopium williamsoni) in the Boundary Area resolved using allelic variation at
microsatellite DNA loci projected using factorial correspondence analysis. The genetic group to the right of the thin vertical grey line
consists of mountain whitefish from Mill Pond and Sullivan Lake as well as three fish sampled from the Pend Oreille River (large orange
symbols with arrows) and one fish sampled from a tributary above Metaline Falls (Sweet Creek, green symbol with arrow right of grey
vertical line). From Small and others (2020) and also includes fish assayed in Small and others (2019a, b).
14 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
alleles. Consequently, the substantial divergence between the
Mill Pond/Sullivan Creek/Sullivan Lake and Boundary Dam/
Albeni Falls mountain whitesh is probably not an artifact of
Mill Pond Dam or Sullivan Lake Dam construction.
The distinction between Mill Pond/Sullivan Creek/
Sullivan Lake mountain whitesh and those from below the
Mill Pond site is also consistent with similar dierences found
in westslope cutthroat trout (for example, Small and others
2017, 2020).
Five sh that were collected below falls, as well as
current or former dam sites were more similar to sh above
these sites suggesting that mountain whitesh can survive
entrainment through natural geographic barriers and dams.
In the case of the single sh inferred to be of upper Sullivan
Creek origin that was collected in the Boundary Dam to
Box Canyon Dam area, the “baseline” sample of sh from
the Sullivan Creek area was likely sucient to support the
inference of downstream movement from Sullivan Creek.
By contrast, the single sh collected in the Boundary Dam
tailrace that was inferred to have originated above the dam
(Small and others, 2019a, b) could not be compared to
mountain whitesh from the mainstem Pend Oreille River
downstream from Boundary Dam or the Salmo River because
there were no baseline samples available for these areas.
Given the limitations of these small sample sizes and their
spatial distributions, a dierent result could arise once baseline
samples from below Boundary Dam are analyzed.
Whiteley and others (2006) used six microsatellite DNA
loci to assay variation among samples of mountain whitesh,
including those collected at the conuence of the Pend Oreille
and Columbia Rivers (PoR-C) and upstream from this area
in the Clark Fork River. The PoR-C sample was more similar
to samples upstream in Canadian portions of the Columbia
River (Beaver Creek, BC, and Kootenay Lake each about
100–110 km upstream from PoR-C) and localities from the
upper (Eagle River, BC) and lower Fraser River (Chilliwack
River, BC, both several hundreds of km away and in a distinct
drainage from PoR-C) than it was to samples from the Clark
Fork drainage (at least 170 km upstream from Boundary
Dam, Z Canyon, and Metaline Falls; Whiteley and others,
2006). High similarity of lower Pend Oreille River mountain
whitesh to sh from the Fraser River compared to sh from
the Clark Fork is inconsistent with high levels of historical
gene ow between the lower Pend Oreille River and areas
upstream.
Ecological Evidence of Connectivity
As for westslope cutthroat trout, water temperature is
a major control on the distribution of mountain whitesh in
stream networks (Isaak and others, 2017a). In a comprehensive
analysis of thermal requirements of 73 sh species in
Wyoming, Mandeville and others (2019) classied species
into ve thermal guilds: cold, cool-cold, cool, cool-warm,
and warm. Cutthroat trout (O. clarkii) was identied as one
of two species (including nonnative brook trout, S. fontinalis)
belonging to the cold-water species guild. Mountain whitesh
was classied as a member of the cool water guild of shes.
Isaak and others (2017a) modeled the presence of mountain
whitesh based on estimated August mean temperatures (Isaak
and others 2017b) in streams located in the upper Columbia
River (including the Pend Oreille River), Snake River, and
Missouri River headwaters. August mean temperatures
associated with estimated probabilities of occurrence of 0.5
for mountain whitesh were 15.3 °C and 9.7 °C for univariate
and multivariate models. It is worth noting that condence
bounds associated with these estimates were relatively wide,
which should be expected with eld-based studies of thermal
associations, as numerous factors interact with temperature
to determine species presence (Mandeville and others, 2019).
Furthermore, associations can vary, depending on descriptors
used to associate sh with thermal regimes (for example,
magnitude, timing, duration, frequency of thermal exposures;
Benjamin and others, 2016). While many uncertainties remain,
available evidence suggests that mountain whitesh are more
likely to be observed using water that is a few degrees warmer
than what can be generally expected of cutthroat trout (Isaak
and others, 2017b). As discussed for westslope cutthroat
trout, summer maximum temperatures within Boundary
Reservoir approach and potentially exceed levels that could
lead to physiological stress in mountain whitesh (Quinn and
others, 2010).
While limited data are available, inferences from
molecular markers (preceding section) indicate little gene
ow and connectivity in the Pend Oreille River downstream
from Boundary Dam for mountain whitesh. Similarly, within
Boundary Reservoir, only six mountain whitesh have been
tagged and tracked, but no individuals were detected being
entrained through Boundary Dam (Hydroacoustics Technology
Incorporated, 2018, 2020, 2021). More individuals would need
to be tracked to draw more denitive conclusions. Movement
studies of mountain whitesh in other systems, along with
knowledge of constraints on connectivity in the Boundary
Reservoir system and Pend Oreille River, can be applied to
draw a few qualied inferences. Direct tracking of individual
movements as well as inferences from otolith microchemistry
of mountain whitesh in other systems indicate seasonal
movements are extensive (for example, up to and potentially
extending over 100 km), complex, and generally limited to
larger streams within riverscapes (for example, Davies and
Thompson, 1976; Northcote and Ennis, 1994; Baxter, 2002;
Benjamin and others, 2014; Boyer and others, 2017). An
improved understanding of spatial and temporally dynamic
habitat features that mountain whitesh may depend on
within Boundary Reservoir could provide useful insights for
maintaining or improving connectivity within the system.
For instance, Davies and Thompson (1976) proposed a host
of proximate (for example, environmental conditions) and
ultimate (for example, growth, survival, reproduction) drivers
of diverse seasonal movements they observed for mountain
whitesh throughout the life cycle.
Part 3. Conclusions and Recommendations for Upstream Passage over Boundary Dam 15
Aside from the isolated distribution of mountain
whitesh in the Sullivan Lake/Creek watershed and the lowest
reach of Sweet Creek, tributary connectivity within Boundary
Reservoir may have little ecological signicance. Given the
lack of access (upstream movement barriers) and limited
suitability (that is, small size) of many tributaries to Boundary
Reservoir, opportunities for mountain whitesh to exploit
seasonally variable conditions within the system are limited.
Evidence from molecular genetic markers indicating lack of
genetic drift in isolated populations of mountain whitesh in
the Sullivan Creek watershed indicates that current and recent
historical population sizes are large enough to overcome the
eects of genetic drift, in spite of their isolation or lack of
connectivity to the Pend Oreille River. Such patterns have
been observed for other salmonid shes upstream from
human-constructed barriers (culverts), where only the smallest
systems show evidence of loss of genetic diversity (Neville
and others 2009). Although some studies have identied
loss of genetic diversity in some instances upstream from
human-constructed and natural passage barriers (for example,
Woord and others, 2005; Neville and others, 2006a, b; Carim
and others, 2016), populations of westslope cutthroat trout
often persist upstream from barriers (Loxterman and others,
2014; Peterson and others, 2014), and at least in the case of
culverts, population persistence times may exceed the lifespan
of culverts (Reagan, 2015). Other structures (for example,
concrete and metal erosion control dams; Morita and others,
2009) may last longer and pose threats to the long-term
persistence of isolated populations.
The importance of upstream connectivity to the Pend
Oreille River above Box Canyon Dam is less clear for
mountain whitesh. As with westslope cutthroat trout, the
ecological importance of connectivity of mountain whitesh
to the Pend Oreille River and associated lakes upstream from
Albeni Falls Dam is unknown. Movements of mountain
whitesh observed in other systems have been extensive
however (for example, Davies and Thompson, 1976;
Northcote and Ennis, 1994; Baxter, 2002; Benjamin and
others, 2014; Boyer and others, 2017), so this is a potential
consideration. As with westslope cutthroat trout and bull trout
(Dunham and others, 2014), there is little possibility, however,
that mountain whitesh were able to move upstream over Z
Canyon and Metaline Falls.
Part 3. Conclusions and
Recommendations for Upstream
Passage over Boundary Dam
Westslope Cutthroat Trout
The historical biogeography of the Pend Oreille River
system, molecular genetic analyses, and contemporary
ecological observations were assembled to draw conclusions
regarding westslope cutthroat trout and the role that Boundary
Dam plays in aecting connectivity for this species. Notably,
the available molecular genetic evidence suggests westslope
cutthroat trout in the Salmo River may: (1) be introgressed
with rainbow trout, and (2) derived from a source outside of
the Salmo basin (King's Lake hatchery strain). We discuss
each of these in turn below.
Introgression between cutthroat trout and rainbow trout
can lead to reduced tness (Strait and others, 2021) and
thus pose a threat to westslope cutthroat trout if one or both
species are introduced outside their natural ranges. There
are likely remnants of native and nonnative rainbow trout
in the Salmo River, owing to historical stocking of rainbow
trout and isolation by downstream dams that have prevented
anadromous (steelhead trout) life history expression (Hagen
and Baxter, 2004). Consequently, westslope cutthroat trout
likely coexist with a mix of native and introduced rainbow
trout in the Salmo River. The potential spread of hybridization
between westslope cutthroat trout and rainbow trout
throughout connected river networks has been documented in
tributaries upstream from the Pend Oreille River (Muhlfeld
and others, 2014), which may be exacerbated by rapid climate
warming observed across the western United States (Isaak and
others, 2016). Similar outcomes are possible if Salmo River
westslope cutthroat trout are connected to native westslope
cutthroat trout in the Pend Oreille River. Given the evidence
from upstream systems, threats posed from hybridization with
nonnative rainbow trout or backcrossing with hybrids merit
serious consideration when evaluating the potential risks
and benets of connecting to isolated native populations of
westslope cutthroat trout in the Pend Oreille River. One option
to facilitate sh passage but protect upstream populations from
introgression would be to remove hybrid individuals passing
over sh passage structures during spawning migrations
between mainstem habitats and tributaries, but this can be a
costly endeavor.
With respect to the hatchery origin (King's Lake
hatchery stock) of westlope cutthroat trout in the Salmo
River,the absence of readily detectable and genetically
unique westslope cutthroat trout in the Salmo River contrasts
strongly with Sullivan Creek immediately upstream, where
westslope cutthroat trout likely represent a native population.
The King's Lake hatchery stock is derived from tributaries
in the Priest Lake watershed (Small and others, 2007) and
can be considered native to the Pend Oreille/Priest Lake
watershed. Consequently, the Salmo River sh could be a
native population and their current similarity to the King's
Lake hatchery strain could be a function of the historical
connectivity between these areas as part of the Pend Oreille
River drainage. Given, however, that the Salmo River
watershed is at the lower end of the Pend Oreille drainage,
it seems unlikely that the genetic similarity between Salmo
River and King's Lake results from historical connectivity
when many other systems closer to the King's Lake hatchery
source populations in the Priest Lake drainage show no
such anity (Small and others, 2007, 2017). Introgression
16 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
following known plantings of King's Lake hatchery sh in and
around the Salmo River seems a more plausible scenario for
the observed genetic structure of Salmo River sh.
Acoustic telemetry in Boundary Reservoir indicates some
tagged westslope cutthroat trout were entrained through the
dam and moved downstream. It is not dicult to imagine this
being the case prior to construction of Boundary Dam when
sh could easily pass downstream over Metaline Falls and
down through Z Canyon to access the Salmo River or other
downstream destinations. Given historical records indicating
that anadromous shes were limited by the falls, any resident
sh moving downstream would likely have been unable to
return upstream and would have been forced to complete their
life cycle downstream. Previous work on bull trout identied
a genetic signal of such interactions, indicating that bull trout
from upstream in the Pend Oreille River or Lake Pend Oreille
may have contributed to populations in the Salmo River but
not vice versa, at least historically (Dunham and others, 2014).
Additionally, our analyses suggest that if sh were passed
upstream from Boundary Dam it is clear that environmental
conditions within Boundary Reservoir are not well-suited to
support westslope cutthroat trout in the warmer months and it
does not appear that sh in the reservoir have easy access to
abundant colder thermal refugia within the system, based on
the presence of natural movement barriers near the mouths of
larger tributaries. In conclusion, the available evidence points
to no strong evolutionary or ecological rationale for passing
westslope cutthroat trout upstream from Boundary Dam.
Mountain Whitefish
Although less is known about mountain whitesh,
relative to westslope cutthroat trout, existing evidence
available within Boundary Reservoir and its tributaries, as
well as information from within and outside of the Pend
Oreille River indicate little support for passing sh upstream
over Boundary Dam. In contrast to westslope cutthroat trout
and bull trout (Dunham and others, 2014), mountain whitesh
within the Pend Oreille River exhibit little population
structure, indicating widespread historical gene ow across the
upper Columbia River downstream from Lake Pend Oreille,
although populations do show structuring at a broader regional
extent (Whiteley and others, 2006). A notable exception
is the uniqueness of mountain whitesh in Sullivan Lake,
which is consistent with the potential existence of a barrier
to upstream movement in the lower reaches of this system
(g. 6). Additional spatial sampling and consideration of a
larger diversity of molecular markers could potentially resolve
additional genetic relationships for mountain whitesh within
the Pend Oreille River and upper Columbia River basin and
potentially shed more light on the importance of upstream
connectivity to the Pend Oreille River and associated lakes
(for example, Dupont and others, 2007). An ecological
study of mountain whitesh within the Pend Oreille River
and Sullivan Creek may reveal important spatial ecological
processes that inuence populations (possibly implied by
patterns of genetic structure, g. 8) within each of those
systems and opportunities for restoration.
Overall Conclusions
Our review of the available evidence for westslope
cutthroat trout and mountain whitesh aligns with the case
for bull trout (Dunham and others, 2014, app. 1), with respect
to the question of whether or not to provide passage of
sh upstream over Boundary Dam. Although each species
exhibits unique characteristics, current evidence indicates that
moving sh upstream over Boundary Dam is not warranted
if the management objective is to maintain natural patterns
of evolutionary and ecological connectivity or to conserve
unique ecological and evolutionary characteristics of extant
local populations of these species in the system. There is
little evidence of natural upstream connectivity due to the
natural, pre-dam inuences of barriers posed by Z Canyon and
Metaline Falls. In the case of bull trout, moving sh upstream
could have adverse consequences for population numbers in
the Salmo River and gene ow between populations upstream
and downstream from Metaline Falls. Such does not seem
to be the case for westslope cutthroat trout, as there is no
denitive evidence of native populations in the Salmo River.
Additional work could, however, be done to verify that native
populations are not present in headwater tributaries of the
Salmo River. Even if such populations were discovered, it is
not clear they were likely connected to populations upstream
from Boundary Dam, as populations of bull trout have been
historically.
Mountain whitesh exhibit much less spatial variability
with respect to their distribution, which is conned to higher
order (larger) streams, and local populations that are weakly
dierentiated relative to westslope cutthroat trout and bull
trout (Whiteley and others 2004; Taylor and others, 2014). An
exception is the presence of unique populations of westslope
cutthroat trout and mountain whitesh in Sullivan Lake–the
largest tributary to Boundary Reservoir. Again, however,
the evidence for Sullivan Creek suggests these populations
have not been inuenced by connectivity with downstream
populations.
Our focus in this report was on the role of Boundary
Dam and the historical connectivity of the two species
considering the upstream migration barriers of Z Canyon
and Metaline Falls in the Pend Oreille River. It is important
to recognize, however, that we did not consider connectivity
farther upstream in the Pend Oreille River such as Box
Canyon Reservoir reaches and the associated systems of
Priest Lake and Lake Pend Oreille. This could be an important
consideration, as the case for bull trout appears to indicate
(app. 1). If connectivity for native shes such as westslope
cutthroat trout, mountain whitesh, and bull trout is a major
References Cited 17
objective within the Pend Oreille River, more work directed at
better understanding connectivity in upstream reaches of the
Pend Oreille River may be warranted.
References Cited
Baxter, C.V., 2002, Fish movement and assemblage dynamics
in a Pacic Northwest riverscape: Corvallis, Oregon State
University, PhD dissertation.
Bellmore, J.R., Pess, G.R., Duda, J.J., O’Connor, J.E., East,
A.E., Foley, M.M., Wilcox, A.C., Major, J.J., Shafroth,
P.B., Morley, S.A. and Magirl, C.S., 2019, Conceptualizing
ecological responses to dam removal—If you remove it,
what's to come?: BioScience, v. 69, no. 1, p. 26–39.
Behnke, R.J., 1992, Native trout of western North America:
American Fisheries Society, Monograph 6, 275 p.
Behnke R.J., 2002. Trout and salmon of North America: New
York, The Free Press, 382 p.
Benjamin, J.R., Brignon, W.R., and Dunham, J.B., 2019,
Decision analysis for the reintroduction of bull trout
into the lower Pend Oreille River, Washington: North
American Journal of Fisheries Management, v. 39, no. 5,
p. 1026-1045.
Benjamin, J.R., Wetzel, L.A., Martens, K.D., Larsen, K.,
and Connolly, P.J., 2014, Spatio-temporal variability
in movement, age, and growth of mountain whitesh
(Prosopium williamsoni) in a river network based upon PIT
tagging and otolith chemistry: Canadian Journal of Fisheries
and Aquatic Sciences, v. 71, no. 1, p. 131–140.
Benjamin, J.R., Heltzel, J.M., Dunham, J.B., Heck, M., and
Banish, N., 2016, Thermal regimes, nonnative trout, and
their inuences on native bull trout in the upper Klamath
River basin, Oregon: Transactions of the American Fisheries
Society, v. 145, no. 6, p. 1318–1330.
Benjamin, J.R., Vidergar, D.T., and Dunham, J.B., 2020,
Thermal heterogeneity, migration, and consequences for
spawning potential of female bull trout in a river–reservoir
system: Ecology and Evolution, v. 10, no. 9, p. 4128–4142.
Boyer, J.K., Guy, C.S., Webb, M.A., Horton, T.B., and
McMahon, T.E., 2017, Reproductive ecology, spawning
behavior, and juvenile distribution of mountain whitesh in
the Madison River, Montana: Transactions of the American
Fisheries Society, v. 146, no. 5, p. 939–954.
Carim, K.J., Eby, L.A., Barfoot, C.A., and Boyer, M.C., 2016,
Consistent loss of genetic diversity in isolated cutthroat
trout populations independent of habitat size and quality:
Conservation Genetics, v. 17, no. 6, p. 1363–1376.
Carim, K.J., Dysthe, J.C., McLellan, H., Young, M.K.,
McKelvey, K.S., and Schwartz, M.K., 2019, Using
environmental DNA sampling to monitor the invasion of
nonnative Esox lucius (northern pike) in the Columbia River
basin, USA: Environmental DNA, v. 1, no. 3, p. 215–226.
Corsi, M.P., Hansen, M.J., Quist, M.C., Schill, D.J., and Dux,
A.M., 2019, Inuences of lake trout (Salvelinus namaycush)
and Mysis diluviana on kokanee (Oncorhynchus nerka)
in Lake Pend Oreille, Idaho: Hydrobiologia, v. 840, no. 1,
p. 351–362.
Davies, R.W., and Thompson, G.W., 1976, Movements of
mountain whitesh (Prosopium williamsoni) in the Sheep
River watershed, Alberta: Journal of the Fisheries Research
Board of Canada, v. 33, no. 11, p. 2395–2401.
DeHaan, P.W., and Bernall, S.R., 2013, Spawning success of
bull trout transported above main-stem Clark Fork River
dams in Idaho and Montana: North American Journal of
Fisheries Management, v. 33, no. 6, p. 1269–1282.
DeHaan, P.W., Bernall, S.R., DosSantos, J.M., Lockard, L.L.,
and Ardren, W.R., 2011, Use of genetic markers to aid in
re-establishing migratory connectivity in a fragmented
metapopulation of bull trout (Salvelinus conuentus):
Canadian Journal of Fisheries and Aquatic Sciences, v. 68,
no. 11, p. 1952–1969.
Dunham, J.B., Taylor, E.B., and Allendorf, F.W., 2014, Bull
trout in the Boundary System—Managing connectivity and
the feasibility of a reintroduction in the lower Pend Oreille
River, northeastern Washington: U.S. Geological Survey
Open-File Report 2014–1229, 28 p., https://doi.org/ 10.3133/
ofr20141229.
Dunham, J.B., Adams, S.B., Schroeter, R.E., and Novinger,
D.C., 2002, Alien invasions in aquatic ecosystems—Toward
an understanding of brook trout invasions and potential
impacts on inland cutthroat trout in western North America:
Reviews in Fish Biology and Fisheries, v. 12, no. 4,
p. 373–391.
DuPont, J.M., Brown, R.S., and Geist, D.R., 2007, Unique
allacustrine migration patterns of a bull trout population in
the Pend Oreille River drainage, Idaho: North American
Journal of Fisheries Management, v. 27, no. 4, p.
1268–1275.
Dux, A.M., Hansen, M.J., Corsi, M.P., Wahl, N.C., Fredericks,
J.P., Corsi, C.E., Schill, D.J., and Horner, N.J., 2019,
Eectiveness of lake trout (Salvelinus namaycush)
suppression in Lake Pend Oreille, Idaho—2006–2016:
Hydrobiologia, v. 840, no. 1, p. 319–333.
18 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Epifanio, J., Haas, G., Pratt, K., Rieman, B., Spruell, P.,
Stockwell, C., Utter, F., and Young, W., 2003, Integrating
conservation genetic considerations into conservation
planning—A case study of bull trout in the Lake Pend
Oreille–Lower Clark Fork River system: Fisheries, v. 28,
no. 8, p. 10–24.
Environmental Science Associates, 2019, Memorandum to
Seattle City Light—Data from temperature attainment
plan monitoring: Portland, Oregon, Environmental Science
Associations.
Falke, J.A., and Fausch, K.D., 2010, From metapopulations
to metacommunities—Linking theory with empirical
observations of the spatial population dynamics of stream
shes, in Community ecology of stream shes—Concepts,
approaches, and techniques: Bethesda, Maryland, American
Fisheries Society Symposium, v. 73, p. 207–233.
Federal Energy Regulatory Commission, 2011, Final
environmental impact statement application for hydropower
license for the Boundary Hydroelectric Project FERC
Project No. 2144–038, Washington and application
for surrender of Hydropower License for the Sullivan
Creek Project FERC Project No. 2225–015, Washington:
Washington, DC, Federal Energy Regulatory Commission,
460 p., accessed November 27, 2021, at http:// www.millpo
nddam.com/ wp- content/ uploads/ 2016/ 04/ FEIS- Boundary-
Sullivan.pdf.
Frantilla, Anne, 2022, Boundary Dam: City of Seattle,
Seattle Municipal Archives, accessed July 3, 2022, at http
s://www.se attle.gov/ cityarchives/ exhibits- and- education/
online- exhibits/ boundary- dam.
Hagen, J., and Baxter, J.S., 2004, Conservation status of
Salmo River rainbow trout—Radio telemetry investigations
of habitat use and abundance: BC Hydro, prepared by John
Hagen and Associates, British Columbia, Canada, accessed
November 27, 2021, at http:// www.sgrc.s elkirk.ca/ bioatlas/
pdf/ Salmo_ Telemetry_ Fina l%20Report PUBLIC.pdf.
Hansen, M.J., Schill, D., Fredericks, J., and Dux, A., 2010,
Salmonid predator-prey dynamics in Lake Pend Oreille,
Idaho, USA: Hydrobiologia, v. 650, no. 1, p. 85–100.
Hydroacoustic Technology Incorporated, 2018, Boundary
Dam sheries biotelemetry study, 2017 Annual Report
HTI-VEMCO Project Ref: P3824: Seattle City Light,
Washington, Agreement Number CC2015-011, 30 p.
Hydroacoustic Technology Incorporated, 2020, Boundary
Dam sheries biotelemetry study, 2019 Annual Report
HTI-VEMCO Project Ref: P3824: Seattle City Light,
Washington, Agreement Number CC2015-011, 31 p.
Hydroacoustic Technology Incorporated, 2021, Boundary
Dam sheries biotelemetry study, 2020 Annual Report
HTI-VEMCO Project Ref: P3824: Seattle City Light,
Washington, Agreement Number CC2015-011, 33 p.
Isaak, D.J., Young, M.K., Nagel, D.E., Horan, D.L., and
Groce, M.C., 2015, The cold‐water climate shield—
Delineating refugia for preserving salmonid shes through
the 21st century: Global Change Biology, v. 21, no. 7,
p. 2540–2553.
Isaak, D.J., Young, M.K., Luce, C.H., Hostetler, S.W.,
Wenger, S.J., Peterson, E.E., Ver Hoef, J.M., Groce,
M.C., Horan, D.L., and Nagel, D.E., 2016, Slow climate
velocities of mountain streams portend their role as refugia
for cold-water biodiversity: Proceedings of the National
Academy of Sciences of the United States of America,
v. 113, no. 16, p. 4374–4379.
Isaak, D.J., Wenger, S.J., and Young, M.K., 2017a, Big
biology meets microclimatology—Dening thermal niches
of ectotherms at landscape scales for conservation planning:
Ecological Applications, v. 27, no. 3, p. 977–990.
Isaak, D.J., Wenger, S.J., Peterson, E.E., Ver Hoef, J.M.,
Nagel, D.E., Luce, C.H., Hostetler, S.W., Dunham, J.B.,
Roper, B.B., Wollrab, S.P., Chandler, G.L., Horan, D.L.,
and Parkes-Payne, S., 2017b, The NorWeST summer stream
temperature model and scenarios for the Western U.S.—A
crowd‐sourced database and new geospatial tools foster
a user community and predict broad climate warming of
rivers and streams: Water Resources Research, v. 53, no. 11,
p. 9181–9205.
Kendall, N.W., McMillan, J.R., Sloat, M.R., Buehrens,
T.W., Quinn, T.P., Pess, G.R., Kuzishchin, K.V., McClure,
M.M., and Zabel, R.W., 2015, Anadromy and residency in
steelhead and rainbow trout (Oncorhynchus mykiss)—A
review of the processes and patterns: Canadian Journal of
Fisheries and Aquatic Sciences, v. 72, no. 3, p. 319–342.
Link, P.K., and Keeley, E.R., 2018, Geologic constraints on
Snake-Columbia River drainage changes and the associated
evolutionary diversication of cutthroat trout over the last
17 million years, in Trotter, P.C., Bisson, P.S., Roper, B.,
and Schultz, L., eds., Cutthroat trout—Evolutionary biology
and taxonomy: Bethesda, Maryland, American Fisheries
Society, p. 53–75.
Loxterman, J.L., Keeley, E.R., and Njoroge, Z.M., 2014,
Evaluating the inuence of stocking history and barriers to
movement on the spatial extent of hybridization between
westslope cutthroat trout and rainbow trout: Canadian
Journal of Fisheries and Aquatic Sciences, v. 71, no. 7,
p. 1050–1058.
References Cited 19
Lukacs, P.M., and Landguth, E., 2020, Boundary hydroelectric
project area Westslope cutthroat trout capture update
2012–2018: Missoula, Montana, University of Montana,
Report prepared for Seattle City Light, 16 p.
Lusardi, R.A., and Moyle, P.B., 2017, Two-way trap and
haul as a conservation strategy for anadromous salmonids:
Fisheries, v. 42, no. 9, p. 478–487.
Mandeville, C.P., Rahel, F.J., Patterson, L.S., and Walters,
A.W., 2019, Integrating sh assemblage data, modeled
stream temperatures, and thermal tolerance metrics to
develop thermal guilds for water temperature regulation—
Wyoming case study: Transactions of the American
Fisheries Society, v. 148, no. 4, p. 739–754.
McPhail, J.D., 2007, The freshwater shes of British
Columbia: Edmonton, Alberta, University of Alberta, 620 p.
McPhail, J.D., and Lindsey, C.C., 1986, Zoogeography of
the freshwater shes of Cascadia (the Columbia system
and rivers north to the Stikine), in Hocutt, C.H., and
Wiley, E.O., eds., The zoogeography of North American
freshwater shes: New York, New York, Wiley Interscience,
p. 615–637.
Miller, B.A., 2006, The phylogeography of Prosopium in
western North America: Provo, Utah, Brigham Young
University, Master's thesis.
Mims, M.C., Day, C.C., Burkhart, J.J., Fuller, M.R., Hinkle, J.,
Bearlin, A., Dunham, J.B., DeHaan, P.W., Holden, Z.A., and
Landguth, E.E., 2019, Simulating demography, genetics,
and spatially explicit processes to inform reintroduction of a
threatened char: Ecosphere, v. 10, no. 2, p. e02589.
Monnot, L., Dunham, J.B., Hoem, T., and Koetsier, P.,
2008, Inuences of body size and environmental factors
on autumn downstream migration of bull trout in the
Boise River, Idaho: North American Journal of Fisheries
Management, v. 28, no. 1, p. 231–240.
Morita, K., Morita, S.H., and Yamamoto, S., 2009, Eects of
habitat fragmentation by damming on salmonid shes—
Lessons from white-spotted charr in Japan: Ecological
Research, v. 24, no. 4, p. 711–722.
Muhlfeld, C.C., Kovach, R.P., Jones, L.A., Al-Chokhachy,
R., Boyer, M.C., Leary, R.F., Lowe, W.H., Luikart, G.,
and Allendorf, F.W., 2014, Invasive hybridization in a
threatened species is accelerated by climate change: Nature
Climate Change, v. 4, no. 7, p. 620–624.
Nellestijn, G., and Ells, B., 2008, A history of the Salmo River
watershed—An overview of events that have had an impact
on our river ecology: Salmo Watershed Streamkeepers
Society, 16 p., accessed November 27, 2021, at h ttp://www.
streamkeep ers.bc.ca/ reports/ Salmo%20River- History
.Impacts.E cology.pdf.
Neraas, L.P., and Spruell, P., 2001, Fragmentation of riverine
systems—The genetic eects of dams on bull trout
(Salvelinus conuentus) in the Clark Fork River system:
Molecular Ecology, v. 10, no. 5, p. 1153–1164.
Neville, H., Dunham, J., and Peacock, M., 2006a, Assessing
connectivity in salmonid shes with DNA microsatellite
markers, in Crooks, K.R., and Sanjayan, M., eds.,
Connectivity conservation: New York, Cambridge
University Press, p. 318–342.
Neville, H.M., Dunham, J.B., and Peacock, M.M., 2006b,
Landscape attributes and life history variability shape
genetic structure of trout populations in a stream network:
Landscape Ecology, v. 21, no. 6, p. 901–916.
Neville, H., Dunham, J., Rosenberger, A., Umek, J., and
Nelson, B., 2009, Inuences of wildre, habitat size, and
connectivity on trout in headwater streams revealed by
patterns of genetic diversity: Transactions of the American
Fisheries Society, v. 138, no. 6, p. 1314–1327.
Northcote, T.G. and Ennis, G.L., 1994, Mountain whitesh
biology and habitat use in relation to compensation and
improvement possibilities: Reviews in Fisheries Science, v.
2, no. 4, p. 347–371.
O’Connor, J.E., Baker, V.R., Waitt, R.B., Smith, L.N.,
Cannon, C.M., George, D.L., and Denlinger, R.P., 2020,
The Missoula and Bonneville oods—A review of ice-age
megaoods in the Columbia River basin: Earth-Science
Reviews, v. 208, p. 1–51.
Peterson, D.P., Rieman, B.E., Horan, D.L., and Young, M.K.,
2014, Patch size but not short‐term isolation inuences
occurrence of westslope cutthroat trout above human‐
made barriers: Ecology of Freshwater Fish, v. 23, no. 4,
p. 556–571.
Pickett, P.J., 2004, Quality assurance project plan—Pend
Oreille River temperature total maximum daily load
technical study: Olympia, Washington, Washington State
Department of Ecology, Environmental Assessment
Program, No. 04-03-109, 49 p.
Pielou, E.C., 1991, After the ice age—The return of life to
glaciated North America: Chicago, Illinois, University of
Chicago Press, 376 p.
Powers, D., 2008, Sullivan Creek RM 0.65 sh barrier
assessment: Olympia, Washington, Waterfall Engineering,
LLC, Prepared for EES Consulting, 8 p., accessed
November 28, 2021, at http s://www.fs .usda.gov/ Internet/
FSE_ DOCUMENTS/ fsbdev3_ 034937.pdf.
20 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Quinn, A.L., Rasmussen, J.B., and Hontela, A., 2010,
Physiological stress response of mountain whitesh
(Prosopium williamsoni) and white sucker (Catostomus
commersoni) sampled along a gradient of temperature and
agrichemicals in the Oldman River, Alberta: Environmental
Biology of Fishes, v. 88, no. 2, p. 119–131.
Reagan, R.E., 2015, Where the stream meets the road—
Prioritizing culvert replacement for sh passage: Corvallis,
Oregon, MS Thesis, Oregon State University, 69 p.
Richter, A., and Kolmes, S.A., 2005, Maximum temperature
limits for Chinook, coho, and chum salmon, and steelhead
trout in the Pacic Northwest: Reviews in Fisheries
Science, v. 13, no. 1, p. 23–49.
Scholz, A., O’Laughlin, K., Geist, D., Uehara, J., Peone, D.,
Fields, L., Kleist, T., Zozaya, I., Peone, T., and Teesatuskie,
K., 1985, Compilation of information on salmon and
steelhead total run size, catch and hydropower related
losses in the upper Columbia River Basin, above Grand
Coulee Dam: Cheney, Washington, Upper Columbia United
Tribes Fisheries Center, Eastern Washington University,
Department of Biology, Fisheries Technical Report, no.
2, 165 p., accessed October 2021 at https: //www.nwco
uncil.org/ reports/ compilation- information- salmon- and-
steelhead- total- run- size- catch- and- hydropower- related.
Seattle City Light (SCL), 2006, Pre-application document
for the Boundary Hydroelectric Project (FERC no. 2144):
Seattle City Light, 692 p., accessed November 28, 2021, at
http s://www.se attle.gov/ light/ Boundary/ Relicense/ docs/ Pre_
Application_ Document_ 04_ 2006.pdf.
Small, M.P., Von Bargen, J., Bowman, C., and Frye, A., 2007,
Cutthroat trout population structure in the lower Pend
Oreille and Priest River subbasins using microsatellite
DNA: Olympia, Washington, Washington Department of
Fish and Wildlife, Conservation Biology Unit, Molecular
Genetics Lab, Report to Seattle City Light.
Small, M.P., Gee, G., and Bearlin, A., 2017, Boundary
hydroelectric project native salmonid genetics program
report—SNP genetic analyses of westslope cutthroat trout
in Sullivan Basin and nearby tributaries of the Pend Oreille
River: Olympia, Washington Department of Fish and
Wildlife, Conservation Biology Unit, Molecular Genetics
Lab, Report prepared for Seattle City Light.
Small, M.P., Kissler, M., and Simmons, R., 2019a, Boundary
hydroelectric project native salmonid genetics program
report—Assign unknown migratory westslope cutthroat
trout collected in Pend Oreille Basin: Olympia, Washington
Department of Fish and Wildlife, Conservation Biology
Unit, Molecular Genetics Lab, Report prepared for Seattle
City Light.
Small, M.P., Terepocki, A., and Simmons, R., 2019b,
Boundary region mountain whitesh genetic draft report:
Olympia, Washington Department of Fish and Wildlife,
Conservation Biology Unit, Molecular Genetics Lab, Report
prepared for Seattle City Light.
Small, M.P., Kissler, M., and Simmons, R., 2020, Boundary
hydroelectric project native salmonid genetics program
report—SNP genetic assignment of westslope cutthroat
trout collected in Boundary Reservoir and tailrace:
Olympia, Washington Department of Fish and Wildlife,
Conservation Biology Unit, Molecular Genetics Lab, Report
prepared for Seattle City Light.
Snyder, M.N., Schumaker, N.H., Dunham, J.B., Keefer, M.L.,
Leinenbach, P., Brookes, A., Palmer, J., Wu, J., Keenan,
D., and Ebersole, J.L., 2020, Assessing contributions of
cold-water refuges to reproductive migration corridor
conditions for adult salmon and steelhead trout in the
Columbia River, USA: Journal of Ecohydraulics, p. 1–13.
Spruell, P., Rieman, B.E., Knudsen, K.L., Utter, F.M., and
Allendorf, F.W., 1999, Genetic population structure within
streams—Microsatellite analysis of bull trout populations:
Ecology Freshwater Fish, v. 8, no. 3, p. 114–121.
Strait, J.T., Eby, L.A., Kovach, R.P., Muhlfeld, C.C., Boyer,
M.C., Amish, S.J., Smith, S., Lowe, W.H. and Luikart, G.,
2021, Hybridization alters growth and migratory life‐history
expression of native trout: Evolutionary applications, v. 14,
no. 3, p. 821–833.
Taylor, E.B., Yau, M.M., and Mattock, A.B., 2014, Population
structure in three species of co-distributed salmonid shes
in the Peace River and tributaries near a major proposed
hydro-electirc development in northeastern British
Columbia, Canada: River Research and Applications, v. 30,
no. 9, p. 1120–1133.
U.S. Fish and Wildlife Service, 2015, Recovery plan for
the conterminous United States population of bull trout
(Salvelinus conuentus): U.S. Fish and Wildlife Service,
Portland District, Oregon, accessed November 28, 2021,
at ht tps://www. federalreg ister.gov/ documents/ 2015/ 09/
30/ 2015- 24670/ endangered- and- threatened- wildlife- and-
plants- recovery- plan- for- the- coterminous- united- states.
Vidergar, D.T., 2000, Population estimates, food habits and
estimates of consumption of selected predatory shes in
Lake Pend Oreille, Idaho: Moscow, Idaho, Master's thesis,
University of Idaho, 109 p.
Warren, M.L., and Burr, B.M., eds., 2014, Freshwater shes
of North America—Petromyzontidae to Catostomidae:
Baltimore, Maryland, Johns Hopkins University Press,
v. 1, 664 p.
References Cited 21
Washington Department of Fish and Wildlife, 2020,
Salmonscape geographic information system: Washington
Department of Fish and Wildlife, accessed November 28,
2021, at htt ps://www.a rcgis.com/ home/ item.html? id= 1e
56a6487185 43ab952e75 9971f086.
Wenger, S.J., Isaak, D.J., Dunham, J.B., Fausch, K.D.,
Luce, C.H., Neville, H.M., Rieman, B.E., Young, M.K.,
Nagel, D.E., Horan, D.L., and Chandler, G.L., 2011,
Role of climate and invasive species in structuring trout
distributions in the interior Columbia River Basin, USA:
Canadian Journal of Fisheries and Aquatic Sciences, v. 68,
no. 6, p. 988–1008.
West Fork Environmental, 2020, Salmonid sampling—
Boundary hydroelectric project area, NE Washington—2019
boundary tributary surveys: Tumwater, Washington, West
Fork Environmental, 21 p.
Whiteley, A.R., Spruell, P., and Allendorf, F.W., 2004,
Ecological and life history characteristics predict population
genetic divergence of two salmonids in the same landscape:
Molecular Ecology, v. 13, no. 12, p. 3675–3688.
Whiteley, A.R., Spruell, P., and Allendorf, F.W., 2006,
Can common species provide valuable information
for conservation?: Molecular Ecology, v. 15, no. 10,
p. 2767–2786.
Woord, J.E., Gresswell, R.E., and Banks, M.A., 2005,
Inuence of barriers to movement on within‐watershed
genetic variation of coastal cutthroat trout: Ecological
Applications, v. 15, no. 2, p. 628–637.
World Waterfall Database, 2022, Homepage: World Waterfall
Database, web, accessed March 3, 2022, at
https://w ww.worldwa terfalldat abase.com/ .
Young, M.K., 1996, Summer movements and habitat use
by Colorado River cutthroat trout (Oncorhynchus clarki
pleuriticus) in small, montane streams: Canadian Journal of
Fisheries and Aquatic Sciences, v. 53, no. 6, p. 1403–1408.
22 Evolutionary and Ecological Connectivity in Washington, Idaho, and British Columbia
Appendix 1. Summary and Update on Connectivity for Bull Trout (Salvelinus
confluentus) in the Pend Oreille River since Dunham and Others (2014)
The dynamic historical and contemporary processes
and features within Lake Pend Oreille and the Pend Oreille
River have likely inuenced evolutionary and ecological
connectivity for native coldwater shes in the system. Of the
native shes present within Lake Pend Oreille and the Pend
Oreille River, bull trout (Salvelinus conuentus) is likely the
most sensitive to changes wrought by humans on the system,
including potential hydrologic changes associated with a
warming climate (Wenger and others, 2011; Isaak and oth-
ers, 2015). Prior to contemporary human inuences on the
system, bull trout likely moved widely throughout the Clark
Fork River, Lake Pend Oreille, and the Pend Oreille River.
Although upstream passage was almost certainly blocked at Z
Canyon and Metaline Falls on the lower Pend Oreille River,
downstream movement of bull trout likely contributed to
populations below these barriers (DuPont and others, 2007;
Dunham and others, 2014).
Genetic evidence indicates this is particularly important
for downstream populations of bull trout in the Salmo River
(Dunham and others, 2014). In fact, human-assisted movement
upstream of these barriers could pose threats to downstream
populations of bull trout, and would not be consistent with the
natural, historical pattern of movement and gene ow in the
system. Although no tributaries to the Pend Oreille River from
the Priest River downstream to the Salmo River are known
to support self-reproducing populations of bull trout, recent
modeling of a potential reintroduction suggest that certain
tributaries (for example Sullivan Creek and Sullivan Lake)
could potentially support local populations if a reintroduction
was implemented (Benjamin and others, 2019; Mims and oth-
ers, 2019). Both of these studies pointed to the importance of
maintaining downstream passage into the Pend Oreille River,
as well as maintaining connectivity within tributaries where
spawning and early rearing of bull trout can be supported.
The case of the Pend Oreille River contrasts with the
major inow to Lake Pend Oreille, the Clark Fork River.
In the Clark Fork River, upstream connectivity has been a
management priority. In that system, bull trout arriving at the
base of Cabinet Gorge Dam upstream of Lake Pend Oreille
are transported to upstream populations if they are genetically
identied as originating from upstream (Neraas and Spruell,
2001; Epifanio and others, 2003; DeHaan and others, 2011).
This program has been eective in connecting individuals
to their natal streams in the absence of their ability to natu-
rally migrate upstream (DeHaan and Bernall, 2013). Similar
success has been reported for other migratory salmonids (for
example, Lusardi and Moyle, 2017).
Existing evidence for the Clark Fork and Pend Oreille
Rivers, as well as smaller tributaries (Spruell and others,
1999) to Lake Pend Oreille, suggests that Lake Pend Oreille
acts as a major source of bull trout moving in and out of these
systems. Evidence also supports a similar dynamic for Priest
Lake and Priest River, the largest tributary to the Pend Oreille
River upstream of the historical upstream passage barriers
of Z Canyon and Metaline Falls. Notably, the movement of
bull trout throughout the system inuences patterns of genetic
connectivity and potentially allows migratory bull trout to
exploit spatial and seasonal variability in prey availability
and availability of refuge from environmental extremes. It is
noteworthy that the current angling record (19 kilograms [kg])
for bull trout was taken from Lake Pend Oreille in 1949. This
record was established as the system was beginning to experi-
ence signicant changes from introductions of a range of spe-
cies intended to support sport sheries, including kokanee (O.
nerka), rainbow trout (O. mykiss), lake trout (S. namaycush),
and Mysid shrimp (Mysis spp.) as forage (Vidergar, 2000).
These species introductions have dramatically altered food
webs within Lake Pend Oreille, and lake trout also prey on
bull trout (Corsi and others, 2019; Dux and others, 2019).
Despite these challenges to bull trout, Lake Pend Oreille still
supports some of the most robust populations of the species
within the Columbia Headwaters Recovery Unit recognized
by the U.S. Fish and Wildlife Service (U.S. Fish and Wildlife
Service, 2015).
In summary, it appears that Lake Pend Oreille, and to a
lesser extent Priest Lake, serve as core habitat for aduivial
bull trout that move in and out of these lakes to areas upstream
in the Clark Fork River and smaller tributaries, or down-
stream through the outlet of the Pend Oreille River, extend-
ing to the Salmo River, and possibly farther during historical
times (Dunham and others, 2014). Connectivity for bull trout
related to these systems, as well as patterns of connectivity
within them, have shaped patterns of genetic variability with
likely consequences for migratory bull trout to express diverse
behaviors and exploit the wide range of conditions available in
the system (for example, Benjamin and others, 2020).
For information about the research in this report, contact
Director, Forest and Rangeland Ecosystem Science Center
777 NW 9th Street, Suite 400
Corvallis, OR 97330
h ttps://www .usgs.gov/ centers/ forest- and- rangeland- ecosystem-
science- center
Manuscript approved on September 15, 2022
Publishing support provided by the U.S. Geological Survey
Science Publishing Network, Tacoma Publishing Service Center

